The narratives of human evolution are oft-told and highly contentious. There are major disagreements in the field about whether human evolution is more like a branching tree or a crooked stick, depending partly on how many species one recognizes. Interpretations of almost every new find will be sure to find opposition among other experts. Disputes often center on diet and habitat, and whether a given animal could walk bipedally or was fully upright. What can we really tell about human evolution from our current understanding of the phylogenetic relations of hominids and the sequence of evolution of their traits?
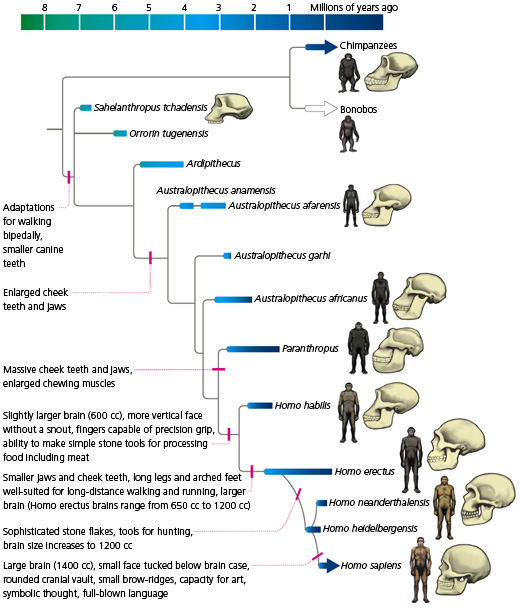
To begin with, let’s take a step back. Although the evolution of hominid features is sometimes put in the framework of “apes vs. humans,” the fact is that humans are apes, just as they are primates and mammals. A glance at the evogram shows why. The other apes — chimp, bonobo, gorilla, orangutan, gibbon — would not form a natural, monophyletic group (i.e., a group that includes all the descendants of a common ancestor) — if humans were excluded. Humans share many traits with other apes, and those other “apes” (i.e., non-human apes) don’t have unique features that set them apart from humans. Humans have some features that are uniquely our own, but so do gorillas, chimps, and the rest. Hominid evolution should not be read as a march to human-ness (even if it often appears that way from narratives of human evolution). Students should be aware that there is not a dichotomy between humans and apes. Humans are a kind of ape.
Virtually all systematists and taxonomists agree that we should only give names to monophyletic groups. However, this evogram shows that this guideline is not always followed. For an example, consider Australopithecus. On the evogram you can see a series of forms, from just after Ardipithecus to just before Homo in the branching order, that are all called Australopithecus. (Even Paranthropus is often considered an australopithecine.) But as these taxa appear on the evogram, “Australopithecus” is not a natural group, because it is not monophyletic: some forms, such as A. africanus, are found to be closer to humans than A. afarensis and others. Beyond afarensis, for example, all other Australopithecus and Homo share “enlarged cheek teeth and jaws,” because they have a more recent common ancestor. Eventually, several of these forms will have to have new genus names if we want to name only monophyletic groups. Students should avoid thinking of “australopithecines” as a natural group with uniquely evolved traits that link its members together and set it apart from Homo. Instead they should focus on the pattern of shared traits among these species and the Homo clade, recognizing that each species in this lineage gains more and more features that are shared by Homo.
In popular fiction and movies, the concept of the wild “ape-man” is often that of a tree-living, vine-swinging throwback like Tarzan. However, the pantheon of hominids is much richer than this, as the evogram shows with forms as different as Paranthropus and Ardipithecus shows. For example, imagine going back in time to the common ancestor of humans and chimps (including bonobos). What did that common ancestor look like? In the Origin of Species Darwin noted that the extinct common ancestor of two living forms should not be expected to look like a perfect intermediate between them. Rather, it could look more like one branch or the other branch, or something else entirely.
Did the common ancestor of humans and chimps conform to the ape-man myth and live in the trees, swinging from vines? To answer this, we have to focus not only on anatomy but on behavior, and we have to do it in a phylogenetic context. Apes such as the gibbon and orangutan, which are more distantly related to humans, are largely arboreal (i.e., tree-living). The more closely related apes such as the gorilla and chimps are relatively terrestrial, although they can still climb trees. The feet of the first hominids have a considerable opposition of the big toe to the others but relatively flat feet, as arboreal apes generally do. But other features of their skeleton, such as the position of the foramen magnum underneath the skull, the vertically shortened and laterally flaring hips, and the larger head of the femur, suggest that they were not just mainly terrestrial but habitually bipedal, unlike their knuckle-walking relatives. Most evidence suggests that the hominid lineage retained some of the anatomical features related to arboreal life and quadrupedal gait even after it had evolved a more terrestrial lifestyle and a bipedal gait. There is no fossil record of these behaviors, but the balance of the available evidence supports the hypothesis that the hominid ancestor was terrestrial and bipedal.
Much discussion in human paleontology surrounds the evolution of a bipedal, upright stance. When and why did this occur? One thing to keep in mind is that “bipedal” and “upright” are not equivalent terms. An animal can be bipedal without having a vertical backbone (think T. rex). It seems clear from the fossil record of hominids that habitual bipedality preceded the evolution of a recurved spine and upright stance. Other changes in the gait, such as how the relatively “splayed” gait of chimps evolved into the gait of humans, who put one foot directly in front of the other, involve studying the hip joint, the femur, and the foot. The famous Laetoli footprints attributed to Australopithecus afarensis are bipedal, but they are still relatively splayed compared to the tracks of living humans.
Another extremely interesting feature in hominid evolution is the degree of sexual dimorphism (i.e., physical differences between the sexes) in different species. Sexual dimorphism is linked to features of sociality and mate competition in many sorts of animals. To understand the evolution of this feature in humans, which have relatively low sexual dimorphism, we need to consider the other apes, in which sexual dimorphism tends to be moderate to high (with exceptions). We don’t have sufficient evidence about Sahelanthropus, Orrorin, and Ardipithecus to understand much about sex differences in these species, but we do know that A. afarensis had relatively high sexual dimorphism: the males were considerably larger than the females. The difference seems to have been less in A. africanus, Paranthropus, and most of the Homo lineage. The evolutionary explanation for A. afarensis‘ dimorphism is not entirely clear. The larger males may have used their size to attract females and/or repel rivals, which would fit with an explanation based on sexual selection. Or the males and females may have been differently sized because they played different roles in their groups, the males hunting and gathering and the females caring for the young. Darwin thought that this differentiation of the sexes may have played a critical role in human evolution, but we simply do not know much about the role of this feature in A. afarensis. Some, all, or none of these functions may have been in play.
We do know that by the time the animals known as Homo evolved, they could make tools, and their hands were well suited for complex manipulations. These features were eventually accompanied by the reduction of the lower face, particularly the jaws and teeth, the recession of the brow, the enlargement of the brain, the evolution of a more erect posture, and the evolution of a limb more adapted for extended walking and running (along with the loss of arboreally oriented features). The evogram shows the hypothesized order of acquisition of these traits. Yet each of the Homo species was unique in its own way, so human evolution should not be seen as a simple linear progression of improvement toward our own present-day form.
Teach your students about human evolution:
- It's all in your head: An investigation of human ancestry, a classroom activity for grades 9-12.
Find additional lessons, activities, videos, and articles that focus on human evolution.