How do complex adaptations evolve when the initial stages of that adaptation do not immediately seem to be adaptive? As mentioned earlier, complex adaptations may arise through intermediate stages that do not initially seem to be adaptive but actually are. In addition, adaptations that evolved in one context may be co-opted for another. However, there are other possibilities:
Lifting functional constraints through duplication
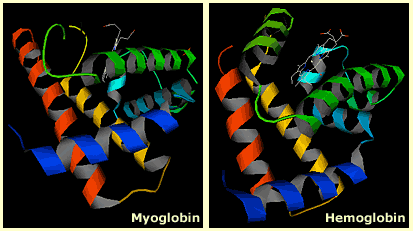
Even when a feature is absolutely necessary for survival it can be modified by natural selection for a different function if it is duplicated. For example, globin is a truly ancient protein. Billions of years old, it was present in the common ancestor of bacteria, plants, animals, and fungi. Globin performed an essential job: binding and carrying oxygen. You might imagine that natural selection would lock globin into that one job; however, through duplication and divergence, different copies of the globin molecule were adapted for different roles. Vertebrates rely on several different globin genes: hemoglobin carries oxygen to body tissues (though a separate globin performs this function in fetuses), myoglobin keeps a reserve supply of oxygen for muscle cells to use, and neuroglobin and cytoglobin do jobs that we don’t yet fully understand. Multiple globin genes are found all across the tree of life. In fact, some globins in deep-sea-dwelling worms are adapted for carrying both oxygen and hydrogen sulfide.
A role for genetic drift
Genetic drift can cause genes that are neutral, or even slightly disadvantageous, to be in high frequencies in small populations. The model that explains how a population might pass through stages of low fitness on its way to high fitness is called the shifting balance theory.
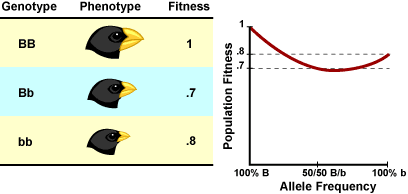
Here’s a simple example of how the shifting balance theory works. Imagine a species of seed-eating bird (like Darwin’s finches) where beak size is controlled by a single gene (of course, in real life, traits like this are controlled by many genes — but the idea is the same for both cases). BB individuals have big beaks, Bb individuals have medium-sized beaks, and bb individuals have small beaks. These birds live in a place where large and small seeds are abundant, but no medium-sized seeds are available. Populations of all big-beaked individuals have a very high average fitness — they can crack open big seeds. Populations of all small-beaked individuals do well (they can manipulate smaller seeds) — but not quite as well as the big-beaked individuals. Medium-beaked individuals have the lowest fitness — they are not particularly good with either big or little seeds (and no medium-sized seeds are available). A graph of these gene frequencies and the population’s resulting fitness levels is shown (below right). This sort of graph is called an adaptive landscape.
Now imagine a small population of all small-beaked individuals (all bb genotypes). They have a high fitness (they are at a local peak), but not as high as a population of big-beaked individuals. Through gene flow some B alleles are introduced to the population. If selection alone were acting, it would weed these alleles out of the population since they would show up in Bb individuals with lower fitness. Under selection alone, the population could never reach the higher BB fitness peak.
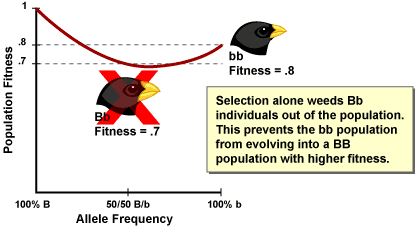
However, since the population is small, drift can be a powerful force. Just by chance, the frequencies of the B alleles increase in the population over several generations (and the population moves into a valley in the adaptive landscape). If the B alleles become frequent enough, the population will begin to have BB individuals with high fitness. As this happens, selection begins to increase the frequency of B (the population moves out of the valley and selection pushes it towards the global fitness peak). Eventually, through the action of genetic drift combined with selection, the population moves from one local peak, through a valley of low fitness, to the global fitness peak.
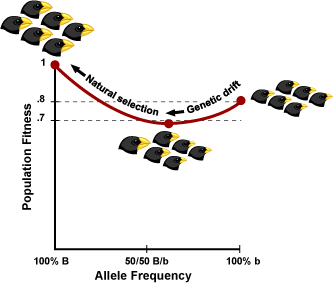
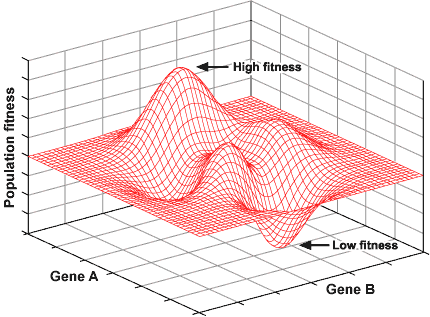
Of course, the above example is a simplified one based on a single locus with two fitness peaks in the adaptive landscape. In the real world, many, many loci affect the fitness of a population and an adaptive landscape may have multiple peaks and valleys. A complex landscape involving just two loci is shown below.