Introduction
Evograms are diagrams that convey information about how a group of organisms and their particular features evolved. The figure below is an evogram. Evograms contain a lot of information, so they cannot be easily digested in a few seconds. However, they are worth understanding because they convey information from several different lines of evidence and are particularly useful in showing students the logic, strength, and testability of evolutionary hypotheses.
What is an evogram?
The evogram below concerns the origin of terrestrial vertebrates. In a separate section we’ll explain more details about this example. Here we just want to show you how evograms are organized.
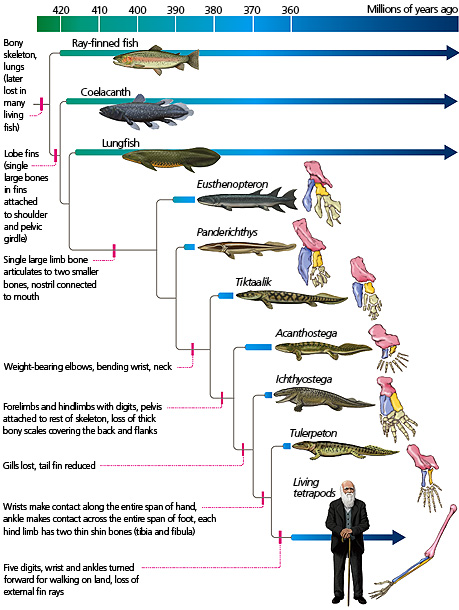
You will see that there are several sorts of information depicted. The centerpiece of the figure is a set of reconstructions of a group of extinct vertebrates (from Eusthenopteron to Tulerpeton) that bridged the transition between water and land in the Devonian period, some 380 million years ago. The blue bars indicate the geologic time span from which fossils of the taxon have been found. These time spans are keyed to the timescale at the top of the page.
On the right side of the image is a set of multi-colored drawings of the limbs of some of those organisms. Although each limb differs in its form from the others, they all have corresponding bones, as indicated by the different colors. From this, students can discern that homologous bones have continuity through evolutionary timescales, and that they changed in shape and number over time.
To the left of the colored drawings is a set of branching lines — an evolutionary tree, or phylogeny, showing the relationships among these animals. It shows that, among the forms pictured here, Tulerpeton is the animal most closely related to today’s tetrapods. Ichthyostega is equally closely related to Tulerpeton and to living tetrapods. Acanthostega is the next most closely related to these three groups. And so on. It’s important to remember that none of these animals is directly ancestral to the other; they are just the closest relatives that we have yet discovered in the fossil record. This is a bit like comparing you, your sibling, your first cousin, and your second cousin: none of them is directly ancestral to you or to the others, but they are successively less closely related to you. You are most closely related to your sibling because you only have to go back one generation to find a common ancestor, whereas you have to go back two generations to find the common ancestor linking you to your cousin. Similarly, Tulerpeton is most closely related to living forms because it shares the most recent common ancestor with these forms compared to the other organisms on the phylogeny.
On various branches of the phylogeny, evolutionary innovations are marked by pink lines. These innovations were inherited by all the lineages descending from the lineage in which the characteristic first evolved. For example, if you examine the diagram, you’ll see that, at some point after Panderichthys split from our ancestors, the lineage leading to tetrapods evolved weight-bearing elbows, bending wrist regions, and a separation between the head and body — a neck. This lineage gave rise to Tiktaalik, Acanthostega, Ichthyostega, Tulerpeton, and living tetrapods, which all inherited these key characteristics.
Phylogenies are not speculations; they are testable hypotheses about evolutionary relationships based on many lines of evidence. To construct a phylogeny, biologists first select the group of organisms that they want to consider. Then they study the characteristics of those organisms — their anatomy,their genetic sequences (if available), and/or the time periods at which they lived. Because evograms can include extinct groups, these phylogenies can be largely based on anatomy. The organisms’ features are coded into a matrix. For example, one feature might be “number of digits” and each animal would be coded as 8, 7, 6, or 5 depending on how many digits it had. Another feature might be “suture in braincase flexible,” which would be coded as “yes” or “no” for each animal. Biologists try to identify as many of these characters as they can. Phylogenies are often based on hundreds of characters.
To help decide which states of these features are ancestral and which evolved later, biologists use “outgroups” — one or more groups that are thought to be less closely related to any of the other organisms on the tree than those organisms are to each other. For example, this evogram uses ray-finned fishes as an outgroup, because we know that they don’t belong to this group of lobe-finned fishes and the earliest tetrapod ancestors. The ray-finned fishes don’t have discrete digits, and neither do coelacanths or lungfishes. This means that the ancestor of this whole group must have had no digits.
The phylogeny is built by taking the matrix of organisms and their features and entering the data into a computer program. These programs use different methods to figure out the most likely set of relationships among the organisms. Commonly, the program is designed to build a tree that requires the fewest evolutionary changes and that groups organisms based on the number and arrangement of newly evolved features. In this case, Tulerpeton and the living forms share the greatest number of these recently evolved features, so they are likely to be each other’s closest relatives. This is how the phylogenetic tree is constructed. Although there are several kinds of information conveyed in evograms, the phylogenetic tree is the most essential kind of information, because without a reliable tree of relationships, we cannot chart the evolutionary change in structures, functions, behaviors, physiology, habitats, and many other features.
Why are evograms important? Evograms show how major evolutionary transitions occurred. The example above shows the evolution of digits and other limb elements. However, we could use the same phylogenetic tree to map different features and ask different questions. We can also use the phylogenies in evograms to test evolutionary hypotheses. Consider, for example, the question of how animals got moveable necks. Most ray-finned fishes don’t have necks (their heads butt squarely against their shoulders), and the most rudimentary necks didn’t move at all. These early necks simply attached the head to the rest of the animal’s body (e.g., as in Tiktaalik and Acanthostega). However, in vertebrates with moveable necks, the first vertebra allows the organism to move its head up and down, and the second allows it to move its head left and right. We could form several hypotheses about how vertebrates evolved this neck. We might suppose that the ability to look right and left evolved first, that the ability to look up and down evolved first, or that they both evolved at once. Similarly, we could form many possible hypotheses about the functions these traits served when they evolved — for example, looking left and right may have helped the organisms see potential prey and predators, and looking up and down may have allowed these new land animals to lower their snouts to pick up food on the ground.
The phylogeny on the evogram helps us test hypotheses about the order in which the traits evolved. For each animal, we can code whether they have a movable first vertebra, a movable second vertebra, both, or neither. Then we map these features on the phylogeny and figure out which evolved first. As it turns out, first the first vertebra acquired its function, and then the second. Of course, this doesn’t tell us what functions these traits served when they first evolved, but sometimes phylogenetic analysis can help us narrow the possibilities.
As we will see, this approach can help us understand how feathers evolved, how mammals got their unique ear, how whales moved back into the ocean and evolved their amazing adaptations, and many other transitions. Using this approach, we can test hypotheses about evolution — as we did for the question of the neck vertebrae. One of the most important lessons we’ve learned from testing hypotheses this way is that structures can evolve new functions over time. For example, the fin-like limbs of animals such as Eusthenopteron and Panderichthys did not provide support on land; these were aquatic animals, and in addition to helping them swim, their fleshy limbs perhaps helped them clamber among the rocks and plants of the shallow waters where they lived. By the time we get to Tulerpeton and Greererpeton, we find limbs in which the individual bones are fewer, stouter, and stronger, and they articulate with each other in a way that can propel the animal on land. So tetrapod limbs evolved in the context of aiding locomotion in aquatic animals and only later evolved to serve the function of locomotion on land. In evolutionary biology, we use the word exaptation to describe a structure that has evolved a new function. Exaptations are common and seem to have been the major way that new functions have evolved.
Feathers are a great example of an exaptation. The first feathers evolved in small carnivorous dinosaurs of the Jurassic period. They were simple, hairlike, filamentous structures that formed a coat all over the animal’s body. They weren’t branched, or downy, or particularly large. They wouldn’t have enabled an animal to fly. But they could have provided warmth, much like down feathers on a living bird do, and probably originally served the function of insulation. In later carnivorous dinosaurs, these feathers evolved more elaborate features. They became branched, and eventually evolved a central stalk with loose filaments around it. They were colored, which could have helped with camouflage or display. Eventually, one of these groups of feathered dinosaurs evolved feathers that formed an effective airfoil and a wing. We call this group the birds. The flight feathers of living birds are exaptations for flight and adaptations for insulation. So to answer a question about the origin of an adaptation (e.g., “what good is half a wing?”), we need to look beyond the feature’s current function to what its original function might have been.
Critics of evolution sometimes charge that scientists don’t have fossils that show the transitions between major evolutionary features and the origins of new adaptations, but that is a misrepresentation. We don’t have fossils of every extinct organism, but we do have fossils from many, many extinct organisms — enough to piece together the stories of some very important evolutionary transitions. And we are learning more every day. All of the examples that we’ll show you here could not have been explained in this detail twenty years ago. The fossils that we’ve discovered in just the last few decades have thrown new light onto these transitions. The longer we search for fossils and study them, the more detailed our knowledge of past life becomes — and the more confident we can be in that knowledge.
Learn more about macroevolution in Evo 101.
Teach your students about macroevolution:
- Visualizing life on Earth: Data interpretation in evolution, a web activity for grades 9-12.
- Interactive investigation: The arthropod story, a web activity for grades 6-12.
Teach your students about evolutionary relationships and phylogenetics:
- What did T. rex taste like?, a web activity for grades 6-12.
Find additional lessons, activities, videos, and articles that focus on phylogenetics.